ここでは、基礎知識編2で軽く解説したケラチンについて、さらに深く解説していきます。
ケラチンの構造
さて、タンパク質は一次構造、二次構造、三次構造、四次構造となるに従って、さまざまな分子間力が働いて、複雑な三次元の立体構造を作り出します。
しかし、ケラチンは同じタンパク質であっても、二次構造のαヘリックス構造を作り出した後は、一般的なタンパク質である三次元の立体構造を作りません。
ケラチン線維は、二次構造のαヘリックス構造の段階で、他の多数のαヘリックスどうしが集まり、「結晶構造」を作り出すことで、非常に強い繊維を作り出しているのです。
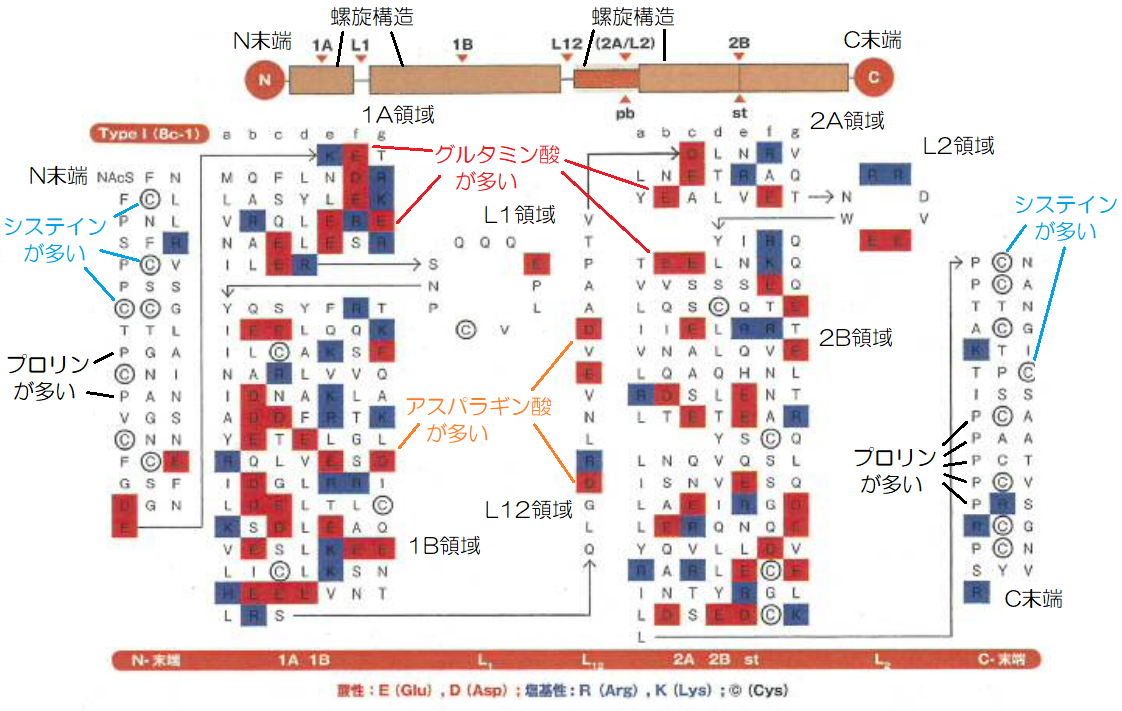 ▲羊毛ケラチンTypeⅠのアミノ酸配列(引用:L.M.Dowling、W.G.rewther and D.A.D.Parry)
▲羊毛ケラチンTypeⅠのアミノ酸配列(引用:L.M.Dowling、W.G.rewther and D.A.D.Parry)
ケラチンは、400個ほどのアミノ酸が結合したポリペプチド鎖からなりますが、αヘリックスはそのすべてで螺旋構造をしている訳ではなく、それは約88%の部分に限られます。
上図は、羊毛のケラチンTypeⅠの全アミノ酸配列で、その上部の模式図はアミノ酸配列の領域、下部のアルファベットの文字列は、アミノ酸の繋がりを現しています。
ケラチンのαヘリックス構造部分は1A、1B、2A、2B領域に分かれ、それを繋ぐ結合部のL1、L12、L2領域、N末端、C末端は螺旋構造が解けた形をしています。
ケラチンTypeⅠは酸性のケラチンであり、アミノ酸配列を見ると、螺旋部分には酸性アミノ酸であるグルタミン酸(E)とアスパラギン酸(D)が多くなっているのが分かります。
一方、ケラチンの特徴であるシステイン(C)は螺旋部分にはほとんどなく、N末端、C末端の方に多く含まれ、同時にその部分にプロリン(P)が多く配列されています。
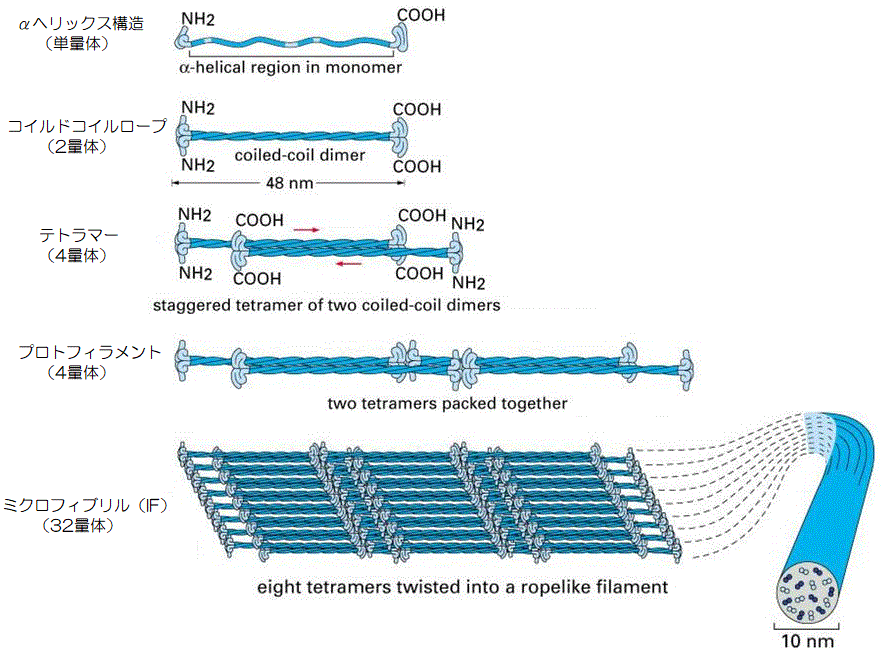
ケラチンは、このようは複雑な形をしていて、この一本のポリペプチド鎖を「ケラチンモノマー(単量体)」と呼び、これが毛髪線維の基本単位になるのです。
コイルドコイルロープの形成
さて、毛髪のケラチンは、その構成アミノ酸に酸性アミノ酸が多いTypeⅠケラチンが9種類、中性や塩基性のアミノ酸が多いTypeⅡケラチンが6種類あります。
そのTypeⅠとTypeⅡの2本のケラチンモノマーがロープのように、今度は左巻きにねじり合い、ダイマー(二量体)を形成し、これを「コイルドコイルロープ」と呼びます。
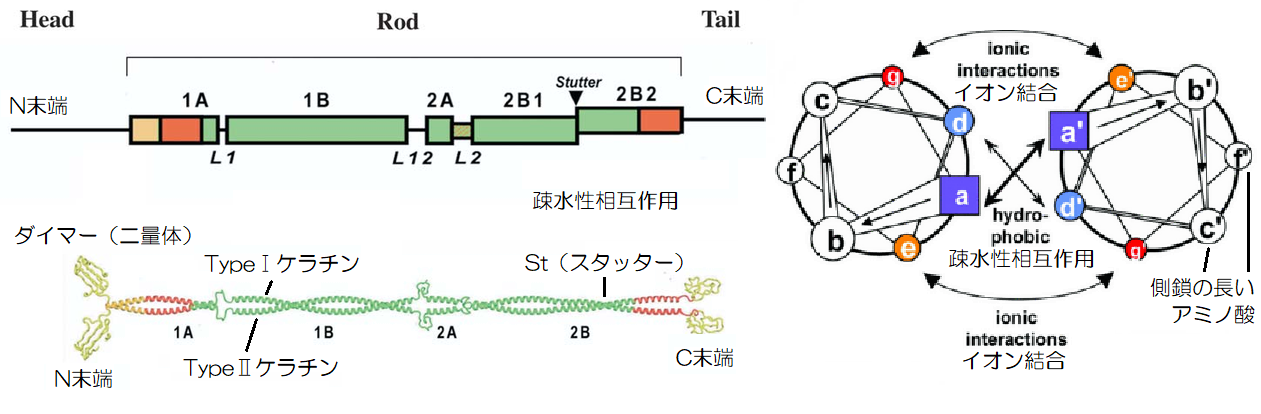 ▲コイルドコイルロープ(ダイマー)の構造(引用:Modified according、Mason et al)
▲コイルドコイルロープ(ダイマー)の構造(引用:Modified according、Mason et al)
そして、ただ単純に2本が絡まり合っている訳ではなく、2A/L2領域はαヘリックス構造が平行になり、2B領域の中間あたりは後戻り(スタッター:st)という螺旋構造が逆転している部分が存在します。
また、αヘリックス構造は、側鎖Rの部分を外側に向けていますが、ダイマーでは内側に側鎖の短いアラニンやロイシンなどの疎水性のアミノ酸が向かい合うように配列されます。
疎水性の分子同士は、水から弾かれる形で集まる性質があり、ダイマーの内側では疎水性相互作用(疎水結合)という力でお互いが引き合っているのです。
また、右図の「g」と「e」のアミノ酸には、プラスの電荷を帯びた酸性アミノ酸、マイナスの電荷を帯びた塩基性アミノ酸などが配列され、互いにイオン結合で引き合っています。
これらの疎水性相互作用、イオン結合などはダイマーの結合を安定化させ、また二重螺旋の平行領域、後戻り領域もダイマーを構成するのには必要な部分であります。
プロトフィラメントの構成
次に、一本のダイマーに対し、もう一本のダイマーがN末端とC末端を逆にして、1/4ずれると、お互いに1B、2B領域が重なり、ここでジスルフィド結合を作って結合します。
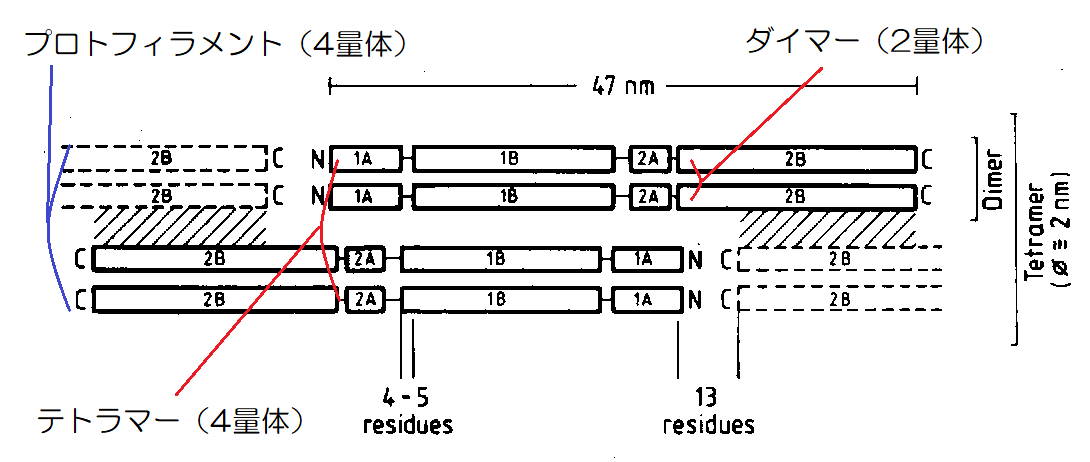 ▲テトラマーの構造
▲テトラマーの構造
この2本のダイマーを「テトラマー(4量体)」と呼び、これが縦に連なって伸びて、繊維状になったものを「プロトフィラメント(4量体)」と呼びます。
そして、このプロトフィラメント(4量体)が次々に集まり、中間径フィラメント(32量体)を構成しますが、その集合方法にはいくつかの説があります。
一つは、プロトフィラメントが8本寄り集まった「ソリッドシリンダーモデル」で、もう一つはプロトフィブリル(8量体)を構成する「ハローシリンダーモデル」です。
中間径フィラメントの構成1
中間径フィラメントの集合方法で、従来から唱えられている説が、「ソリッドシリンダーモデル」です。
 ▲ソリッドシリンダーモデルのIF鎖構造(引用:Garland Science,New York)
▲ソリッドシリンダーモデルのIF鎖構造(引用:Garland Science,New York)
この説は、プロトフィラメント(4量体)が8本集まって、それがロープのように捻じれるように束ねられることで、「ミクロフィブリル(32量体)」を構成します。
このミクロフィブリルの構造が、細胞骨格である中間径フィラメント(Intermediate Filament)であり、「IF(鎖)」などと略され、直径7nm、長さ1μmほどになります。
つまり、中間径フィラメントは断面でみると、ケラチンモノマー(単量体)が32本、またダイマー(2量体)単位では16本集まった形になるのです。
中間径フィラメントの構成2
一方、新しい説である「ハローシリンダーモデル」では、プロトフィラメント(4量体)が2本、右巻き螺旋構造をすることで、「プロトフィブリル(8量体)」を作り出します。
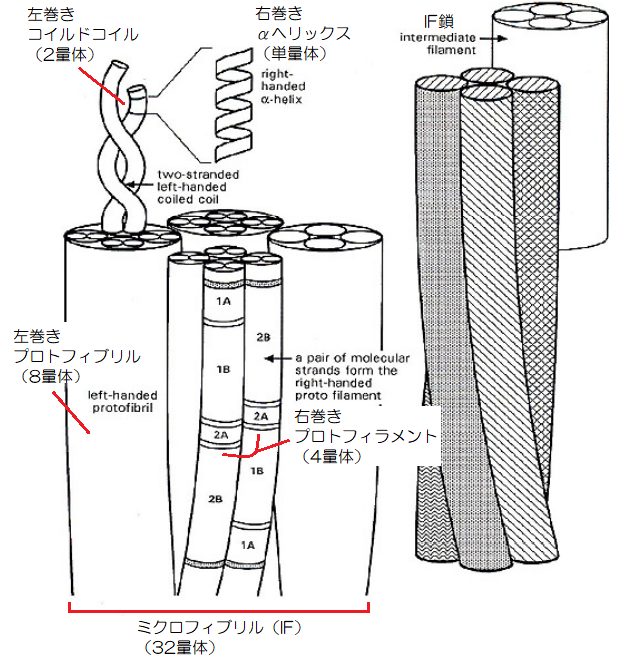 ▲ハローシリンダーモデルのIF鎖構造(引用:Technical University of Cluj-Napoca)
▲ハローシリンダーモデルのIF鎖構造(引用:Technical University of Cluj-Napoca)
そして、そのプロトフィブリル(8量体)が、4本集まり、左巻き螺旋構造をしてミクロフィブリル(32量体)を構成し、直径が7~10nmほどになります。
このモデルでは、αヘリックス構造が右巻き、コイルドコイルが左巻き、プロトフィラメントが右巻き、プロトフィブリルが左巻きと、ケラチンが集まるに従って螺旋の向きが逆になっているのが分かると思います。
この構造により、螺旋構造が崩れにくくなって繊維の強靭さを作り出すと考えられますが、中央にはソリッドシリンダーモデルにはない空間が生まれるのが特徴です。
ケラチン結合タンパク質(KAP)
さて、ケラチン線維は、中間径フィラメントの単位まで集まってくると、周りには別のケラチン線維が次々と結合してきます。
それが、球状タンパク質である「ケラチン結合タンパク質(Keratin associated protein)」、通称KAP(もしくはIFAP)です。
KAPは、アミノ酸が80個~160個ほど連なった短めのポリペプチド鎖が不規則に折れ曲がり、全体として球体の構造をしています。
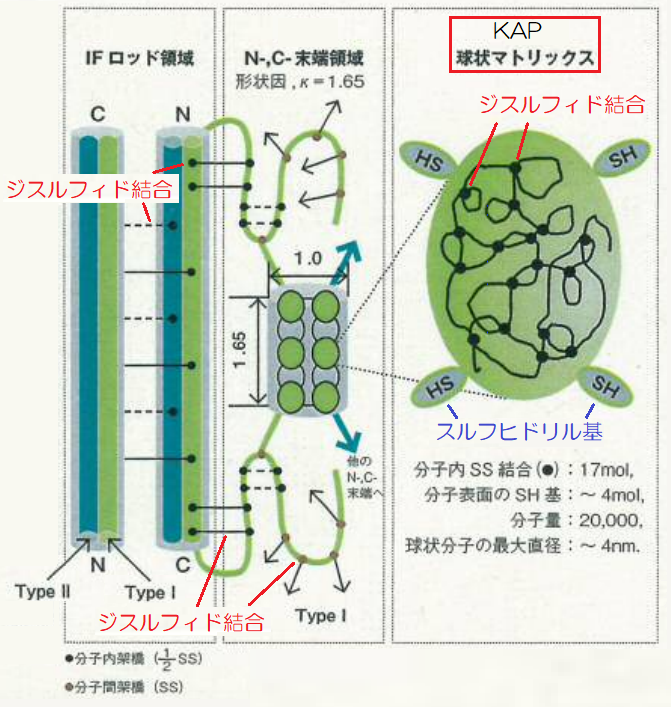 ▲ハローシリンダーモデルのケラチン構造
▲ハローシリンダーモデルのケラチン構造
KAPのアミノ酸配列は、(-システイン-X-Y-Z-システイン-)というように、システインが4残基ごとに現れたβベンド構造と呼ばれる構造をし、分子間でジスルフィド結合を形成しています。
そして、システインの中で、4つのSH基(スルフヒドリル基)は外側に向いていて、他のKAPやIF鎖などと強固にジスルフィド結合をして安定化しています。
このKAPは、IF鎖より弾力性が高いため、髪を折っても、ヒビが入ったり割れたりしないなど、毛幹に柔軟性をもたらします。
また、KAPは通常のタンパク質とは異なり、疎水基を内側、親水基を外側に向けているため、水分を非常に吸収しやすい性質があります。
くせ毛に多く含まれるオルトコルテックスは、IF分子よりこのKAPの割合の方が多いため、水分を吸収して膨張しやすく、湿度が高いとくせが強くなってしまうのです。
IF+KAPの構造
さて、中間径フィラメントのほどけているN末端とC末端は、システインを多く含みますが、どのような構造になっているかは最近まで解明されていませんでした。
しかし、最近の研究では、N末端とC末端はαヘリックス構造の外側に出て、網目構造をなし、互いにジスルフィド結合をしていると推測されています。
そして、その網目構造部分に、システインを多く含む球状タンパク質のKAPがジスルフィド結合を始めとした結合で、安定化されていると考えられるのです。
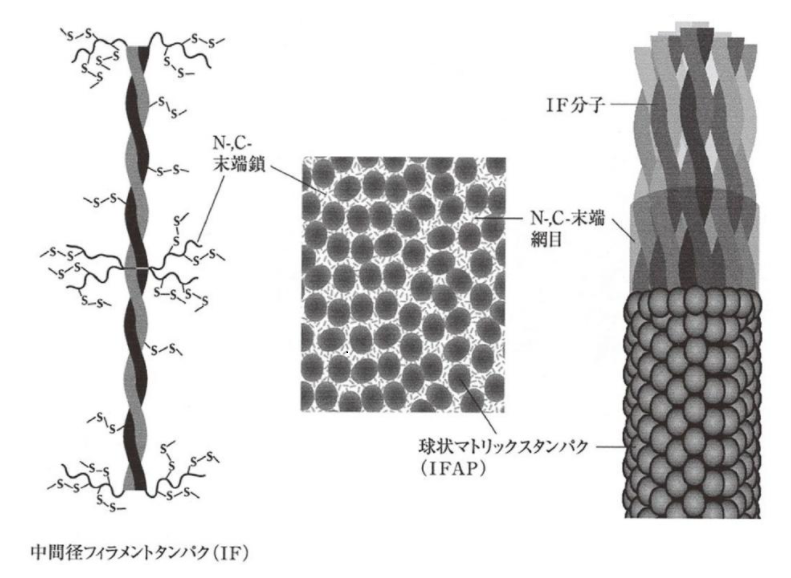 ▲IFとKAPの構造
▲IFとKAPの構造
IFの網目構造とKAPの結合にはジスルフィド結合の他、水素結合、イオン結合、疎水性相互作用などの複数の作用が働いていて、IFを取り巻くようにKAPが配置されています。
この強靭なIFフィラメントと、弾力性のあるKAPという複合組織が、毛髪に「強靭さ」と「しなやかさ」という、相反する力を与えているのです。
中間径フィラメントの六角晶構造
さて、周囲をKAPに取り囲まれた中間径フィラメントは、下左図のように、中央に1本、周囲に6本集まって、横断面で見ると鉛筆のような六角晶の構造を形成します。
 ▲IFの六角晶構造(引用:Academic Press,New York)
▲IFの六角晶構造(引用:Academic Press,New York)
上図の中央は、パラコルテックス部分の横断面の電子顕微鏡画像で、六角形の構造を作ったIF鎖が無数に集まっているのが確認できます。
ここでは、IF鎖どうしが周囲のKAPのシステインを介して、ジスルフィド結合で強固に繋がっていると考えられますが、詳細には解明されていません。
一方、上右図のオルトコルテックスの画像は、パラコルテックスとは異なり、六角晶構造がかなり潰れてしまい、指紋状になっているのが確認できます。
オルトコルテックスのIF鎖がこのように潰れて見えるのには原因があります。
マクロフィブリルの形成
マクロフィブリルは、上記のIF鎖の六角晶の構造が無数に集まったものであり、幅が約300nm、長さが約10μm(100,000nm)という細長い大きさになります。
下図が、カールした毛幹外側のオルトコルテックス、内側のパラコルテックスのマクロフィブリルの横断面(上)と縦断面(下)の電子顕微鏡画像です。
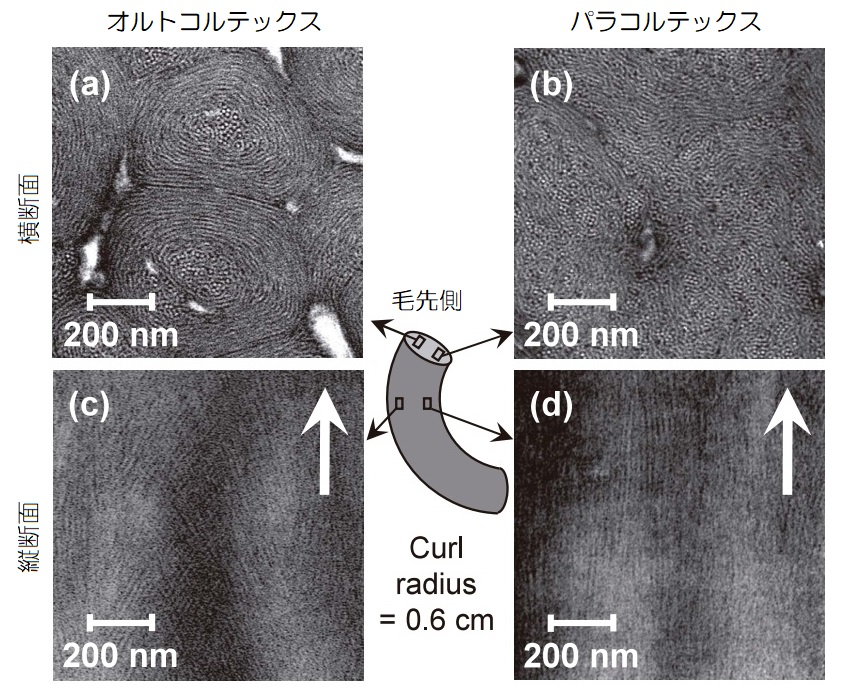 ▲毛幹マクロフィブリルの電子顕微鏡画像(引用:花王、ビューティケア研究センター)
▲毛幹マクロフィブリルの電子顕微鏡画像(引用:花王、ビューティケア研究センター)
右側、パラコルテックスの横断面はIF鎖の白い点状が綺麗に確認でき、縦断面を見ると、IF鎖が毛軸方向に対して平行して配列しているのが確認できます。
一方、左側のオルトコルテックスの横断面は、マクロフィブリルが同心円状の指紋のような構造をし、中央は白い点状に見えますが、外側になるほど潰れているように見えます。
そして、縦断面を見ると、パラコルテックスとは異なり、IF鎖が毛軸方向に対して、斜めに交差して配列している様子が観察できます。
これは、オルトコルテックスのIF鎖が捻じれて螺旋状に配列しているため、横断面を見るとIF鎖が潰れ、縦断面では交差しているように見えているためなのです。
次へ▶【くせ毛考察編1】ネット上にある「くせ毛になる理由」のウソと真実