ここでは、前章に続いて「毛包」の構造についてより詳しく解説します。
結合組織性毛包(CTF)
結合組織性毛包(結合組織性根鞘・真皮鞘)は、真皮と連続し、毛包の一番外側を取り囲んでいて、外側の「外線維膜」と内側の「内線維膜」に分けられます。
結合組織性毛包
・外側:外線維膜(縦走線維層・外縦走層):膠原繊維が縦に走る
・内側:内線維膜(輪走線維層・内輪状層):膠原繊維が円形に走る
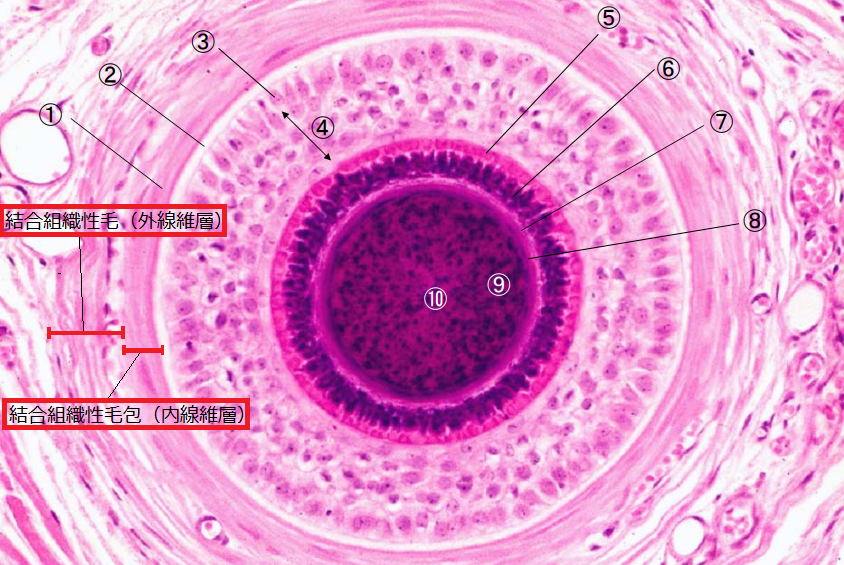 ▲毛包の断面図(引用:神戸大学/改)
▲毛包の断面図(引用:神戸大学/改)
結合組織性毛包は、真皮と連続する組織のため、真皮と同様に膠原線維が走り、弾性線維と脂肪細胞も少量見られますが、細胞成分はあまり見られません。
外側の外線維膜では膠原線維が縦に走り、それに沿うように毛細血管も縦に流れていて、少量の脂肪細胞を含み、縦走線維層や外縦走層などとも呼ばれます。
一方、内側の内線維膜では、膠原線維が毛包を取り巻くように円形状に走り、それに沿って外線維膜より豊富な毛細血管が円形に走り、神経線維の走行も確認できます。
これらの豊富な毛細血管から内側の硝子膜を通して、外毛根鞘細胞や内毛根鞘細胞へと酸素や栄養素が送られます。
また、結合組織性毛包のバルジ領域部の膠原線維には立毛筋が結合し、寒冷時などに毛を立ち上がらせる働きがあります。
硝子膜(GM)
硝子膜は、表皮の基底膜から連続した1層の薄い層で、結合組織性毛包と外毛根鞘とを隔てています。
硝子膜はその名の通り、透明度が高く、非常に丈夫な層で、内側は棘のようになり、外毛根鞘に食い込んでいます。
毛包が退行期を迎えると、アポトーシスといって、外毛根鞘や内毛根鞘などの毛包は次第に消滅しますが、硝子膜は消失することがありません。
そのため、毛包の退行と共に凝縮されて肥厚し、毛根部に透明のゼリー状になって付着するため、毛が抜けた時にはっきりと観察することができるのです。
外毛根鞘(ORS)
外毛根鞘(外根鞘)は、硝子膜の内側にあり、表皮の基底細胞や有棘細胞から連続する上皮系成分の層で、外毛根鞘細胞からなります。
外毛根鞘
・外側:円柱細胞層(表皮の基底層に相当)1層
・中央:多形細胞層(表皮の有棘層に相当)数層
・内側:コンパニオン層(表皮の淡明層に相当)1層
 ▲毛包の断面図(引用:science photo gallery/改)
▲毛包の断面図(引用:science photo gallery/改)
外毛根鞘は、硝子膜に対して垂直に並んだ円柱細胞層(基底層)1層と、その内側にある数層の多形細胞層、そして一番内側の透明なコンパニオン層の3層からなります。
円柱細胞層は表皮の基底層、多形細胞層は有棘層から連続する層で、機能もほぼ同じであり、表皮と同様に細胞間隙と細胞間橋の構造が見られます。
この2層に、結合組織性毛包からの組織液が流れ込み、細胞分裂を繰り返すことで、毛包を肥大させ、毛包を皮下組織の奥へと陥入させていきます。
また、多形細胞層は表皮の有棘細胞と同様、内毛根鞘に近づくにつれ扁平な構造に変化し、その内側は掌に見られる淡明層と似た、非常に薄いコンパニオン層が確認できます。
コンパニオン層は小さな毛包では見られませんが、表皮の顆粒層と同様、細胞間は密着結合により強固に繋がり、毛孔部から入ってきた水分や異物の侵入を防ぐ役割を果たします。
また、内側のヘンレ層とデスモソームにより結合し、毛幹の成長により内毛根鞘と共に上昇しますが、外側の多形細胞層は移動しないため、2層の摩擦を干渉する役割を担います。
外毛根鞘の特徴
外毛根鞘を縦断面でみると、毛球部では1~2層と薄く、毛乳頭の頂点を超えると多層化し、毛隆起部では7~12層程度になり、さらに上方の毛包峡部では、再び薄くなります。
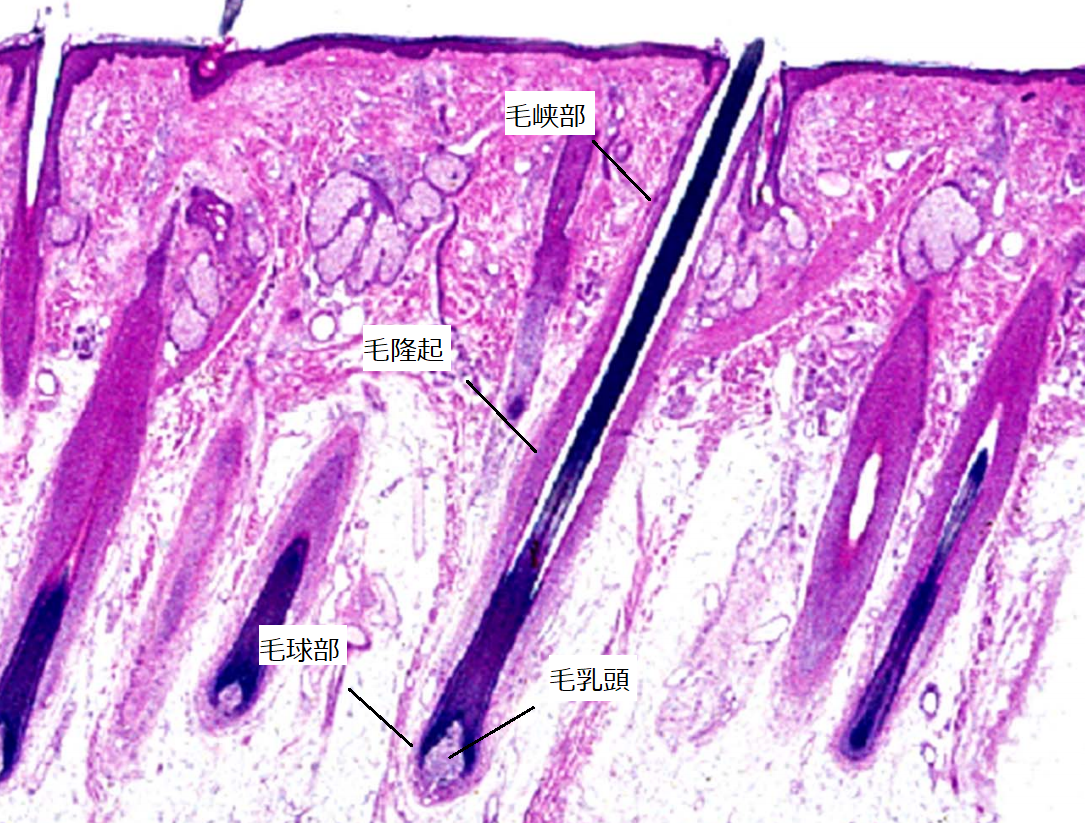 ▲毛包の縦断面(引用:神戸大学/改)
▲毛包の縦断面(引用:神戸大学/改)
最も多層化したバルジ領域部では、毛包幹細胞、色素幹細胞が存在し、17型コラーゲンにより硝子膜に係留されていて、成長初期に毛母細胞や色素細胞へと分化します。
外毛根鞘の多くは角化しませんが、内毛根鞘から離れる毛包峡部では、ケラトヒアリン顆粒を形成せずに角化する外毛根鞘角化が見られ、毛幹を毛包に繋ぎとめる働きがあります。
また、外毛根鞘細胞はエストロゲン受容体(ER)を有し、エストロゲンが外毛根鞘の発達に関与していることがうかがえます。
内毛根鞘(IRS)
内毛根鞘(内根鞘)は外毛根鞘の内側にあり、毛幹と直接接する部分であり、外側からヘンレ層、ハックスレー層、鞘小皮(根鞘小皮)の3層に分けられます。
内毛根鞘
・外側:ヘンレ層1層
・中央:ハックスレー層1~3層
・内側:鞘小皮1層
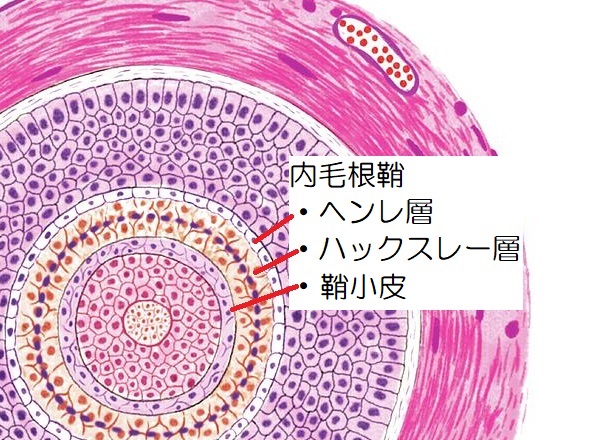 ▲毛包の横断面(引用:science photo gallery/改)
▲毛包の横断面(引用:science photo gallery/改)
内毛根鞘は、表皮の顆粒層のケラトヒアリン顆粒や、角層のケラチン線維のように、3層ともトリコヒアリン顆粒とケラチン線維を含んでいます。
トリコヒアリン顆粒は、表皮の角化細胞のケラトヒアリン顆粒に相当し、ケラチン線維を細胞内で束ねる働きがあり、その役割を果たすと消失します。
内毛根鞘は毛幹と同じように角化し、毛幹の成長と共に内毛根鞘も上昇していき、やがて毛幹から離れて毛孔から排出され、皮脂膜と混じり合ってフケになります。
内毛根鞘の3層と角化
内毛根鞘の最外層であるヘンレ層は、低立方の1層の細胞であり、ハックスレー層は1~3層の丸みを帯びた有核細胞で、内側の鞘小皮は1層の角化した扁平な鱗状の層です。
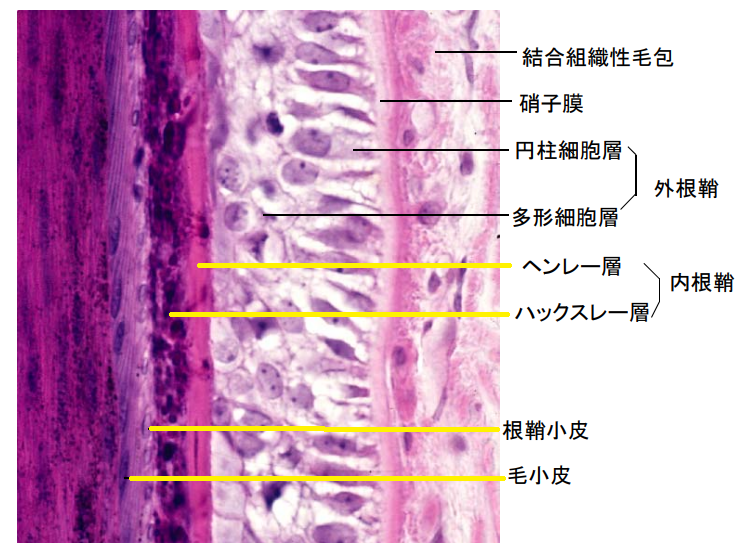 ▲毛包の縦断面(引用:神戸大学/改)
▲毛包の縦断面(引用:神戸大学/改)
ヘンレ層は、毛乳頭の頂点を超えた辺りで、トリコヒアリン顆粒とケラチン線維を蓄積し、毛包の中で最も初期に角化し、その後トリコヒアリン顆粒は消失します。
ハックスレー層も、トリコヒアリン顆粒とケラチン線維が見られ、内毛根鞘の中でも角化は最も遅く、毛包の1/4程度になってようやく角化が完成します。
鞘小皮は、毛幹のキューティクルと同じような鱗状の細胞で、毛幹の内側の下方に向けてケラチン線維を形成し、内側のキューティクル層は外側の上方へ向けて鱗状に並んでいます。
そのため、お互いがフック状に噛み合い、この構造が毛幹と毛包を繋ぎとめる働きを持ちますが、細胞接着の構造は持たないため、接着力自体は弱くなっています。
鞘小皮は、ヘンレ層の次にトリコヒアリン顆粒によりケラチン線維を満たして角化が完了し、その後トリコヒアリン顆粒は消失して、キューティクル層から離れます。
内毛根鞘の角化とくせ毛
内毛根鞘は、毛包の1/4程度まで来ると、角化が全て完了して薄くなり、毛幹から離れて隙間が生じます。
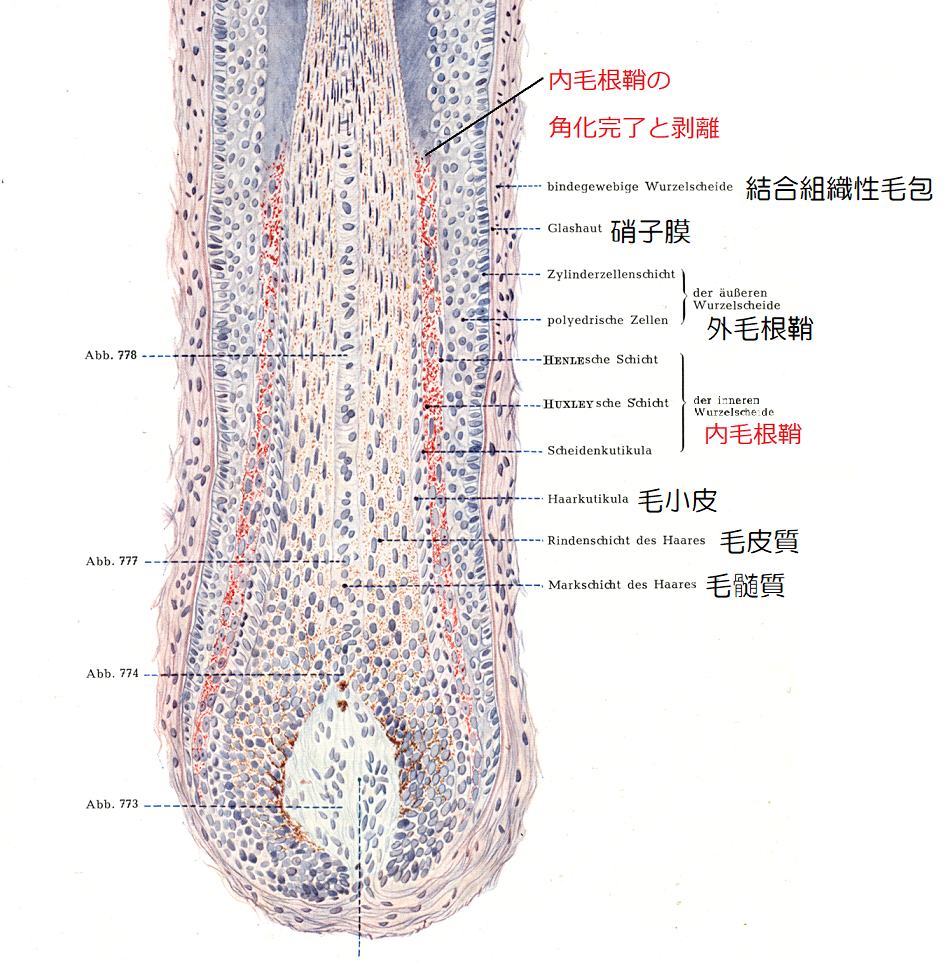 ▲毛包の縦断面(引用:Rauber Kopsch/改)
▲毛包の縦断面(引用:Rauber Kopsch/改)
角化を終えた内毛根鞘は、やがて剥がれ落ち、毛幹の成長と共に毛孔部へと押し出され、その上方に開く脂腺から分泌される皮脂膜と混じり合ってフケを生成します。
内毛根鞘は、毛幹より先に角化が完了するため、毛幹を支える働きがあると共に、まだ角化していない柔らかい毛幹の断面形状を誘導する「鋳型」としての性質があります。
つまり、内毛根鞘の形状が毛幹の形を決めるため、その断面が真円だと毛幹は直毛になり、楕円のように歪んでいると、毛幹の断面もゆがむことになり、くせ毛を発症するのです。
そのため、この部分にKRT74遺伝子やL1PH遺伝子、TCHH遺伝子などの異常があると、内毛根鞘が真円形状を構築できず、縮毛症や乏毛症を発症し、くせ毛になります。
また、内毛根鞘細胞はアンドロゲン受容体を発現しているため、思春期に分泌が増えるテストステロンの増加が、内毛根鞘の形状に影響を与えることが考えられます。